Max Zinke
§ 00.1 — Thesis
Making molecular
machinery accessible.
§ 00.2 — About
I am a scientist who loves building things. PhD and postdoc in molecular biophysics with a strong focus on structural biology (Institut Pasteur, FMP Berlin); currently Scientist at Bachem in Switzerland. In my free time I make scientific visualizations, build NMR tools, and occasionally stumble into web development/design.
§ 01 — What I do
Three pillars.
-
01.1
NMR Spectroscopy
Biomolecular NMR — method development at the intersection of classical spectroscopy and modern computation.
-
01.2
Scientific Visualization
Blender renders of proteins and mechanisms, recently for EvolutionaryScale.
-
01.3
Scientific Software
Full-stack, end-to-end. ML-based tools for NMR data analysis.
§ 02 — Selected work
Figures.

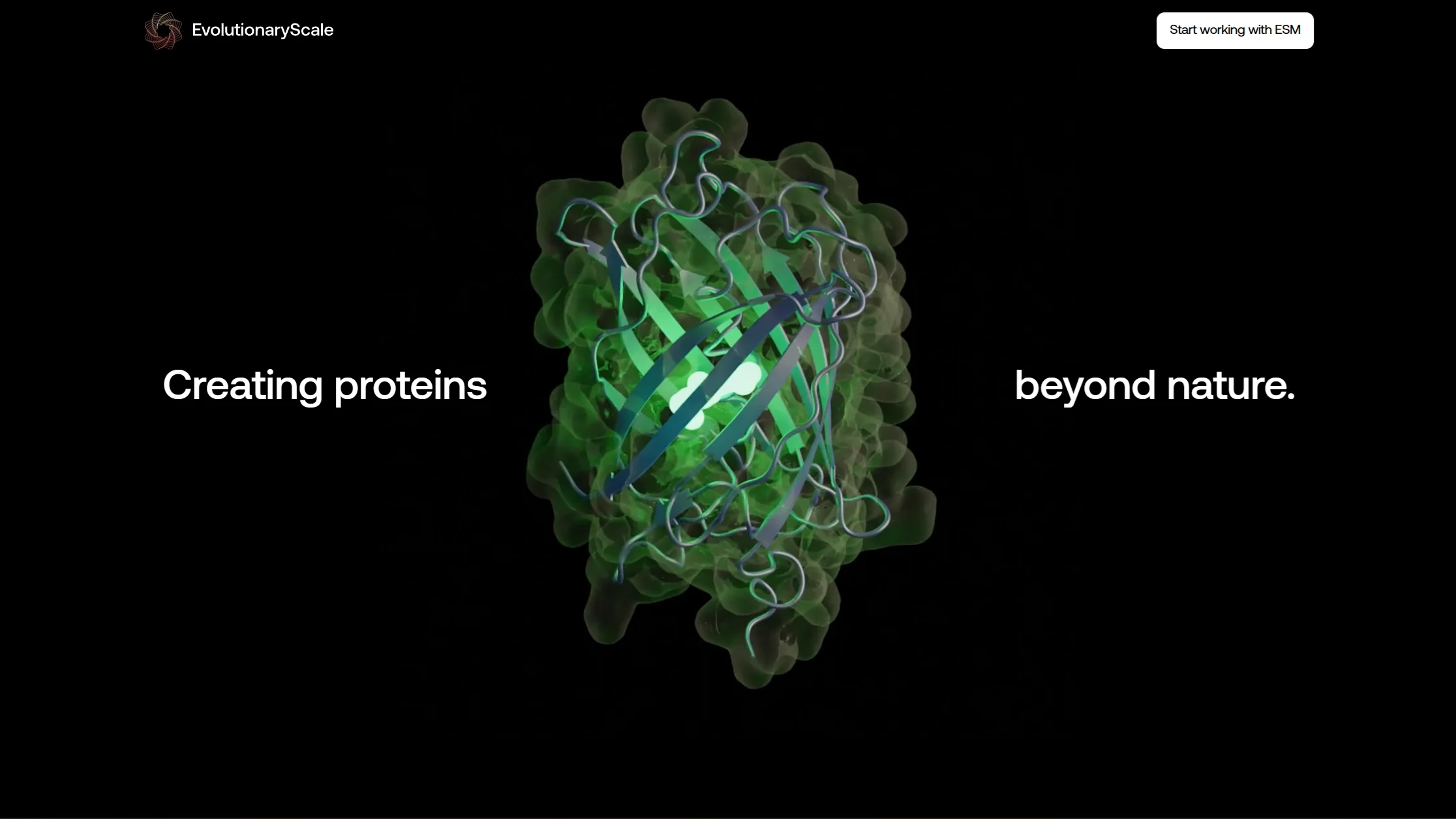
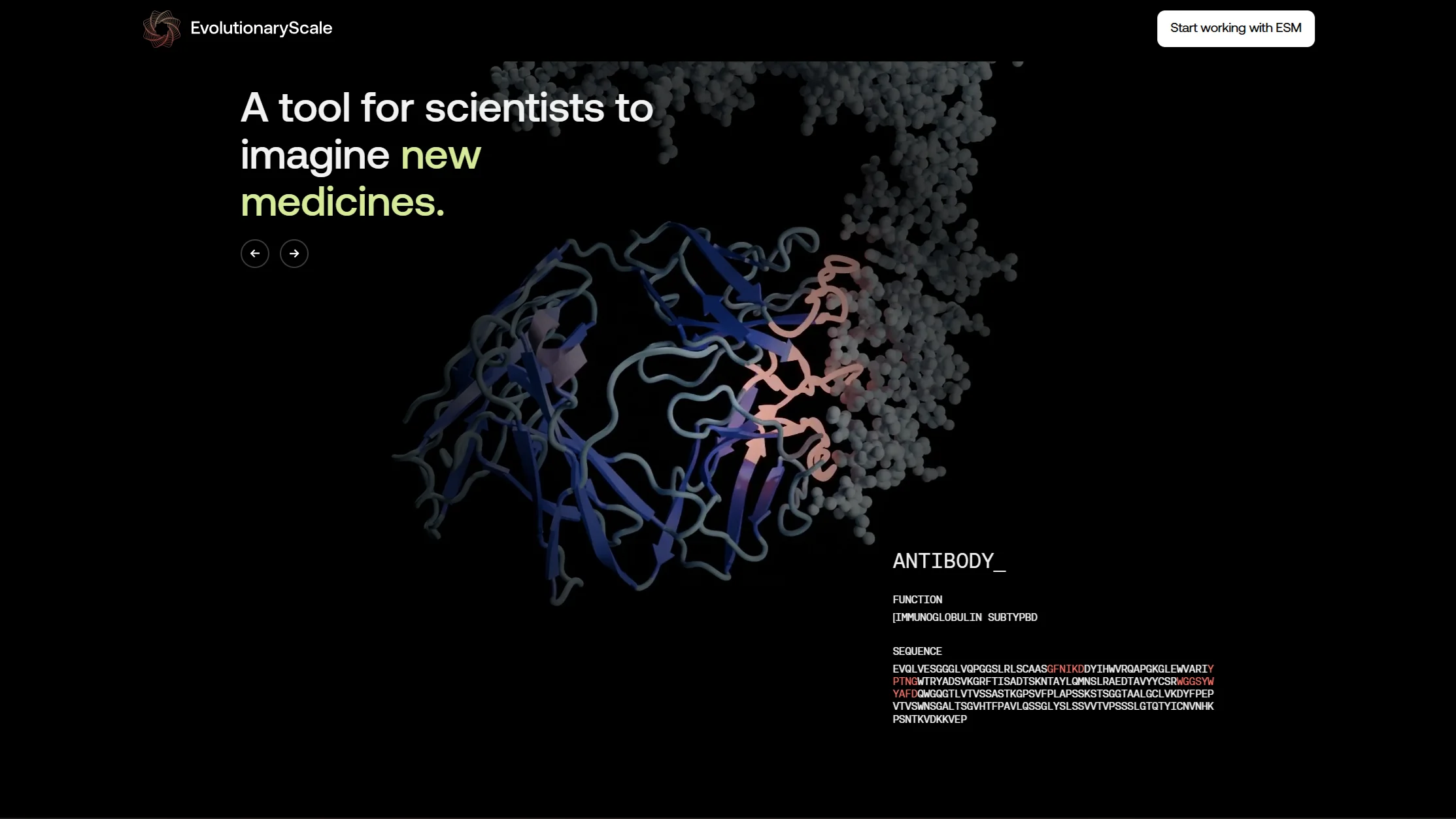
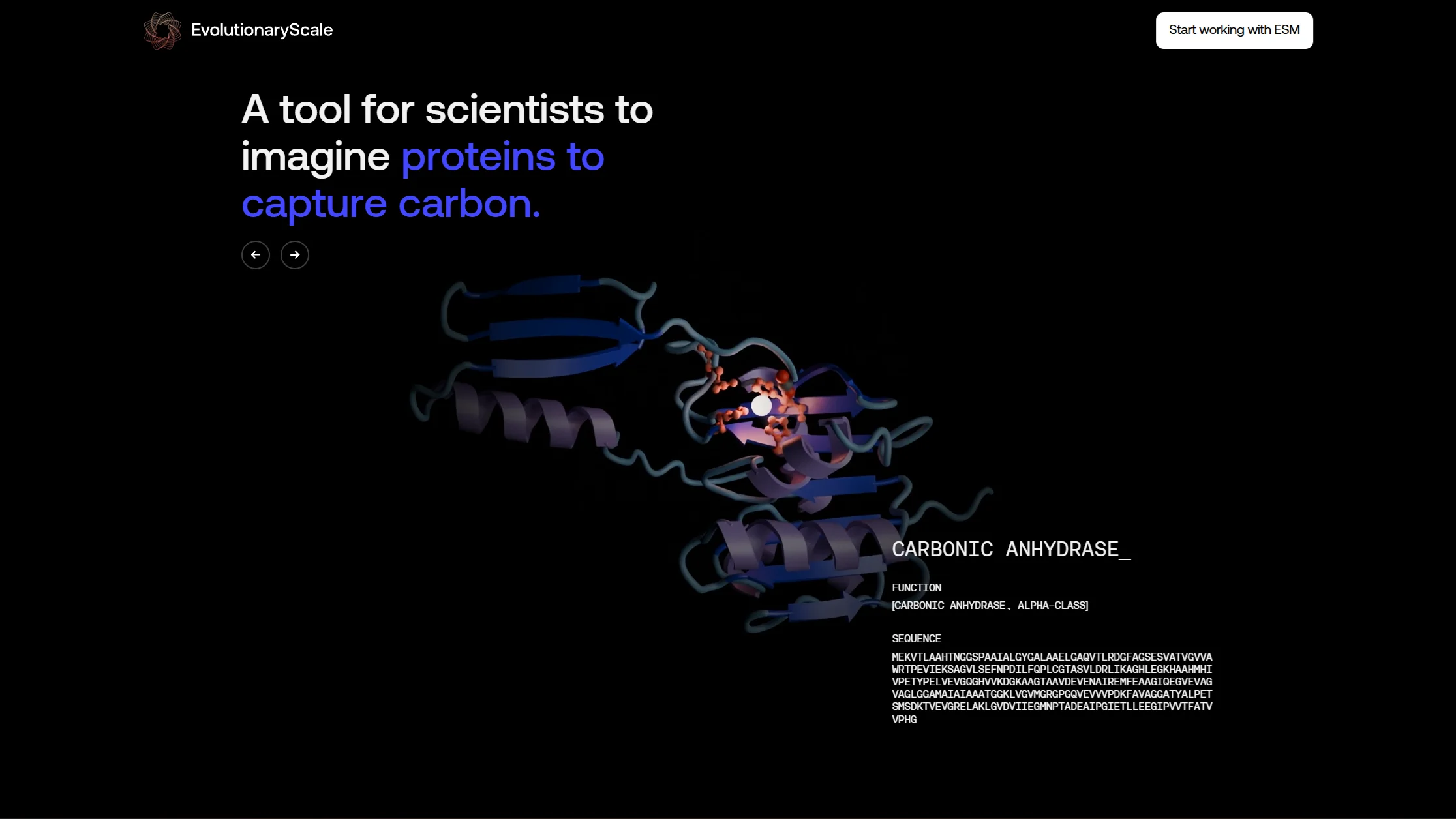
These sit on the landing page of EvolutionaryScale, the lab behind ESM3. The brief was to show protein-language-model-generated structures as something investigable and beautiful, not as another glowing molecule on a dark gradient. See for yourself.
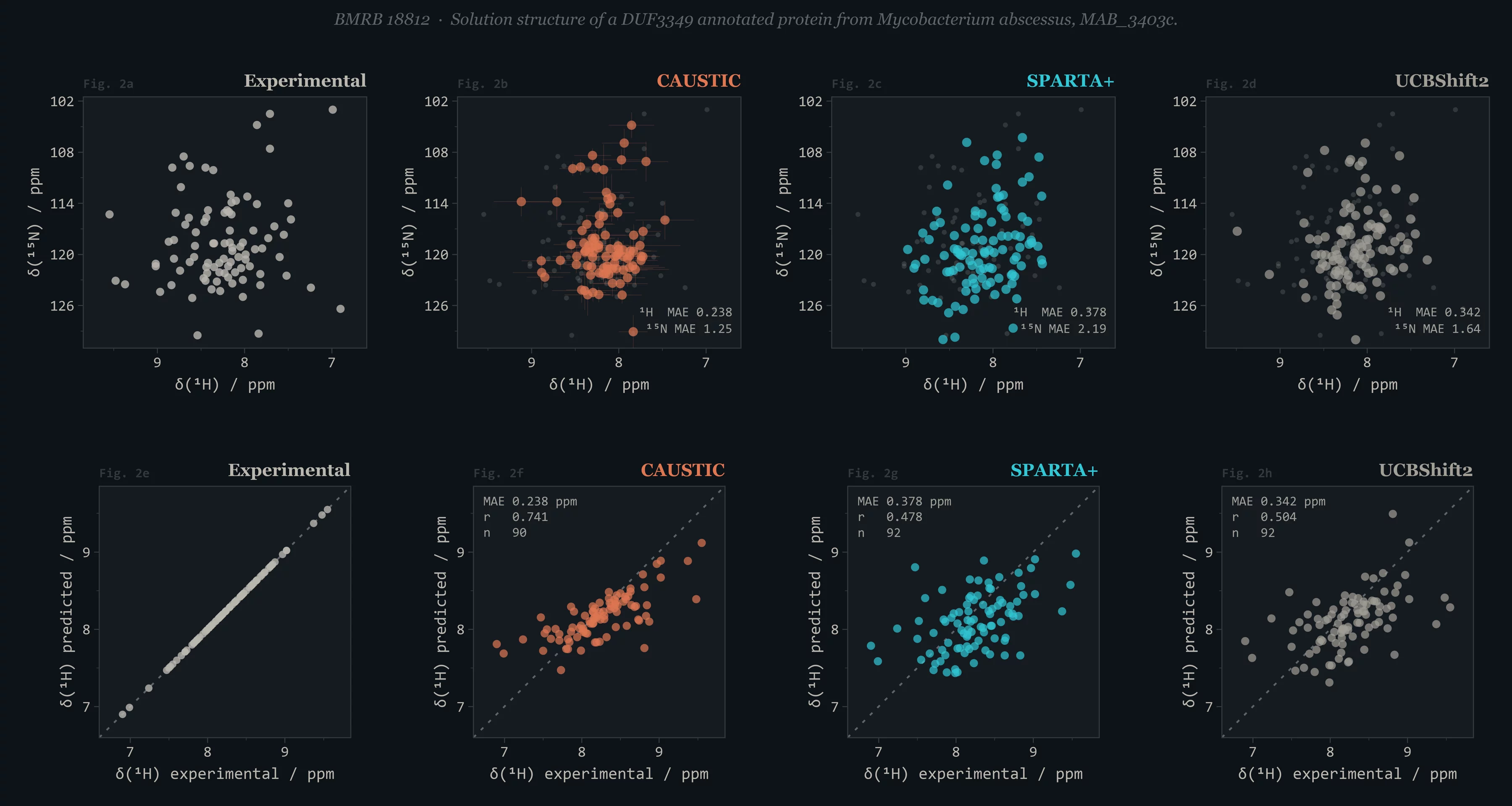
A 739K-parameter equivariant PaiNN graph neural network trained on 3,433 BMRB-linked experimental structures. Predicts all six backbone nuclei (¹H, ¹Hα, ¹⁵N, ¹³Cα, ¹³Cβ, ¹³C') from any PDB, mmCIF, or AlphaFold structure — with calibrated uncertainties. Beats SPARTA+, LEGOLAS, and UCBShift2 on every nucleus across a 735-protein, homology-separated test set (e.g. ¹Hα MAE 0.156 ppm vs 0.217 / 0.239 / 0.265; ¹³Cα MAE 0.72 ppm vs 0.88 / 1.01 / 1.46). Try it in your browser.
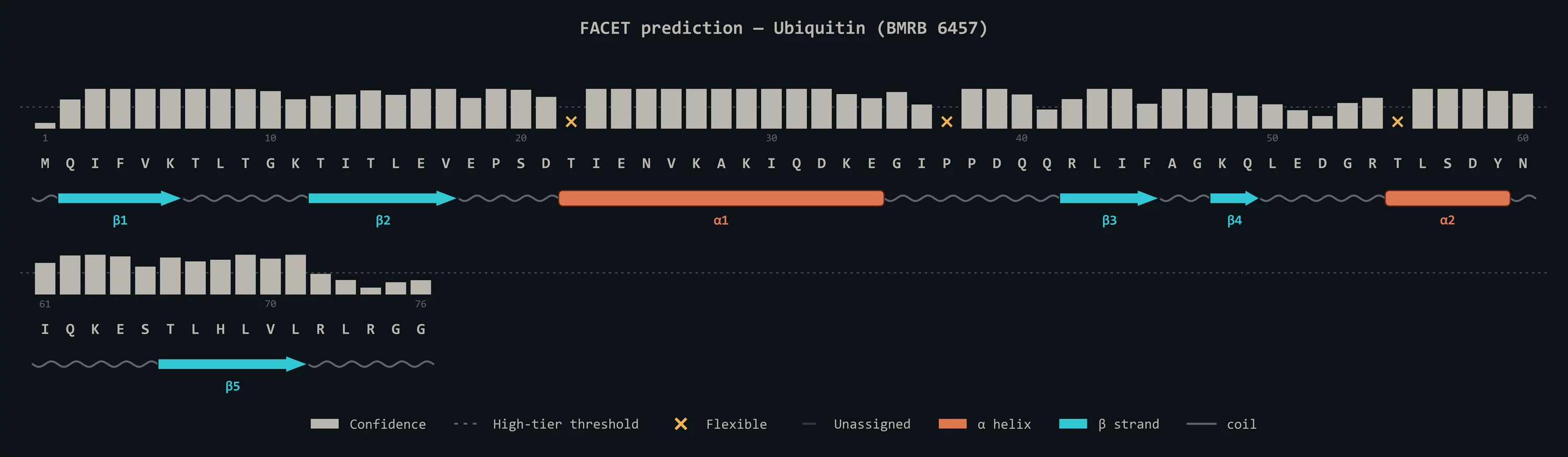
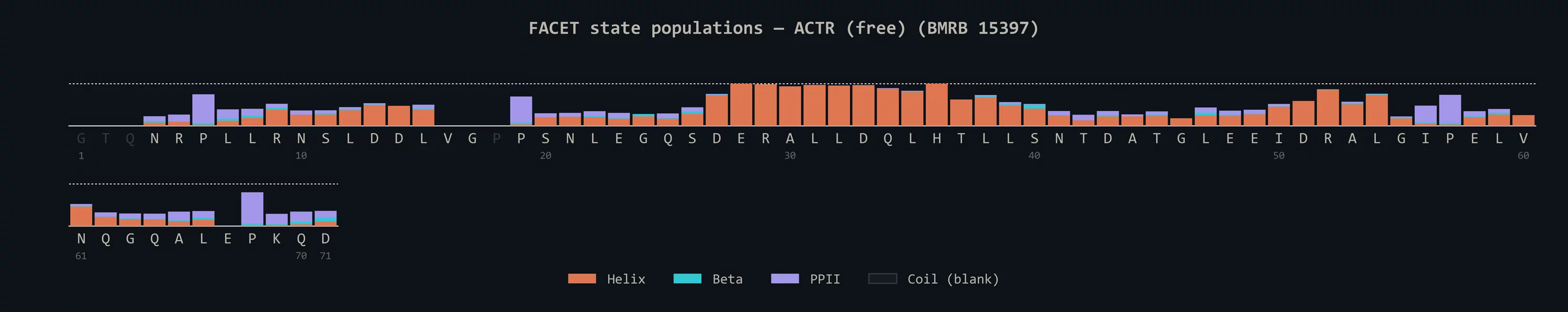
A 1.3M-parameter retrieval-augmented transformer with k-NN lookup over a 220K-residue reference index. Predicts (φ, ψ) with calibrated confidence tiers, secondary structure for folded proteins, and α / β / PPII state populations for IDPs — directly from ¹H, ¹Hα, ¹⁵N, ¹³Cα, ¹³Cβ, ¹³C' chemical shifts, no structure required. (a) Ubiquitin (BMRB 6457): the canonical β-grasp topology fully recovered. (b) ACTR free state (BMRB 15397, fold-on-binding IDP): the transient α-helix at residues 26–37 — the segment that locks in upon NCBD binding — cleanly resolved. Beats TALOS-N by 1.26° (median Δθ) in the high-confidence "Strong" tier on a 55,036-residue, homology-separated test set. Try it in your browser.
An animation of the mechanism I solved during my postdoc at Institut Pasteur: the conformational switch of ExbD that drives active nutrient uptake in Gram-negative bacteria, and the catalytic role of the peptidoglycan layer. The Ton system harnesses the proton motive force to open outer-membrane transporters — our work showed that ExbD's periplasmic domain is a dynamic dimer switching between open and closed states, and that this switch triggers a disorder-to-order transition of TonB. First-author paper in Nature Communications.
A live, shipped product in a completely unrelated field: Counter-Strike 2 case-hardened skin patterns, reverse-engineered, rendered, and searched at scale. It is on this page for one reason — it is proof I build end-to-end products that real people use.
§ 03 — Contact
Reach me.
- linkedin.com/in/maximilian-zinke
- publications
- 17 papers, 2016–2024 →